

Deleted member 43589
Well-Known Member
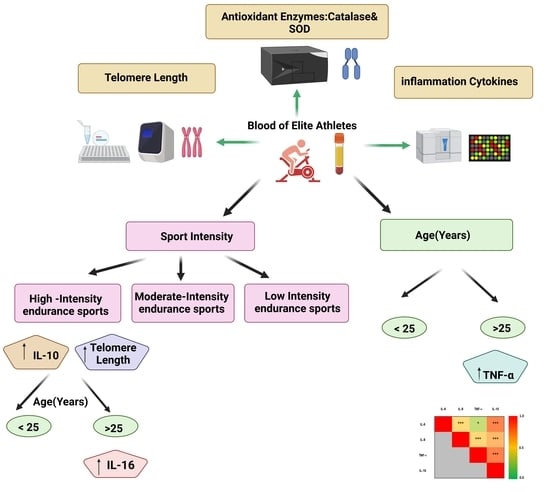
Age and Sport Intensity-Dependent Changes in Cytokines and Telomere Length in Elite Athletes
Exercise-associated immune response plays a crucial role in the aging process. The aim of this study is to investigate the effect of sport intensity on cytokine levels, oxidative stress markers and telomere length in aging elite athletes. In this study, 80 blood samples from consenting elite...
Sellami M, Al-muraikhy S, Al-Jaber H, Al-Amri H, Al-Mansoori L, Mazloum NA, Donati F, Botre F, Elrayess MA. Age and Sport Intensity-Dependent Changes in Cytokines and Telomere Length in Elite Athletes. Antioxidants. 2021; 10(7):1035. Age and Sport Intensity-Dependent Changes in Cytokines and Telomere Length in Elite Athletes
Abstract
Exercise-associated immune response plays a crucial role in the aging process. The aim of this study is to investigate the effect of sport intensity on cytokine levels, oxidative stress markers and telomere length in aging elite athletes. In this study, 80 blood samples from consenting elite athletes were collected for anti-doping analysis at an anti-doping laboratory in Italy (FMSI). Participants were divided into three groups according to their sport intensity: low-intensity skills and power sports (LI, n = 18); moderate-intensity mixed soccer players (MI, n = 31); and high-intensity endurance sports (HI, n = 31). Participants were also divided into two age groups: less than 25 (n = 45) and above 25 years old (n = 35). Serum levels of 10 pro and anti-inflammatory cytokines and two antioxidant enzymes were compared in age and sport intensity groups and telomere lengths were measured in their respective blood samples. Tumor necrosis factor-alpha (TNF-α) was the only cytokine showing significantly higher concentration in older athletes, regardless of sport intensity. Interleukin (IL)-10 increased significantly in HI regardless of age group, whereas IL-6 concentration was higher in the older HI athletes. IL-8 showed a significant interaction with sport intensity in different age groups. Overall, significant positive correlations among levels of IL-6, IL-10, IL-8 and TNF-α were identified. The antioxidant catalase activity was positively correlated with levels of TNF-α. Telomere length increased significantly with sport intensity, especially in the younger group. HI had longer telomeres and higher levels of pro- and anti-inflammatory cytokines, suggesting less aging in HI compared to low and moderate counterparts in association with heightened immune response. Investigation of the functional significance of these associations on the health and performance of elite athletes is warranted
So it appears that the intensity of exercise pays a very big part in the ability to live longer. The more intensively the discipline you practice, the slower your cells age. This was show by the increased length of telomers and the higher levels of pro- and anti-inflammatory cytokines. Note intensity also pays a big part in the immune system.



 Mark Burnley
Mark Burnley












